مکانیسم انقباض عضله اسکلتی
در حالت طبیعى، فیبرهاى عضله اسکلتى از طریق فیبرهاى عصبى میلین دار قطور (آلفا- موتونورون ها) تحریک مى شوند.این فیبرهاى عصبى در محلى موسوم به صفحه محرکه یا محل تماس عصبى- عضلانى(Neuromuscular Junction) با فیبرهاى عضله اسکلتى تماس برقرار مى کنند(تصویر زیر).
 به استثناى دو درصد فیبرهاى عضلانى، فقط یک صفحه محرکه براى هر فیبر عضلانى وجود دارد که در نزدیکى مرکز فیبر قرار گرفته است.بنابراین، پتانسیل عمل از وسط فیبر به سوى دو انتهاى آن انتشار مى یابد.این نوع گسترش پتانسیل عمل از این نظر اهمیت دارد که موجب انقباض تقریبا همزمان تمام سارکومرهاى عضله مى شود به طورى که این سارکومرها مى توانند به جاى انقباض انفرادى،به طور دسته جمعى و با هم منقبض شوند.
به استثناى دو درصد فیبرهاى عضلانى، فقط یک صفحه محرکه براى هر فیبر عضلانى وجود دارد که در نزدیکى مرکز فیبر قرار گرفته است.بنابراین، پتانسیل عمل از وسط فیبر به سوى دو انتهاى آن انتشار مى یابد.این نوع گسترش پتانسیل عمل از این نظر اهمیت دارد که موجب انقباض تقریبا همزمان تمام سارکومرهاى عضله مى شود به طورى که این سارکومرها مى توانند به جاى انقباض انفرادى،به طور دسته جمعى و با هم منقبض شوند.
فیزیولوژی ناحیه تماس عصبی-عضلانی(Neuromuscular Junction)
فیبر عصبى در انتهاى خود منشعب شده و مجموعه اى از ترمینال هاى عصبى موسوم به صفحه انتهایى(end plate) تشکیل مى دهد،که در یک فرورفتگى از سطح فیبر عضلانى قرار مى گیرد اما به طور کامل در خارج غشاء پلاسمایى فیبر عضلانى قرار دارد (تصویر فوق)
فرورفتگى غشاء فیبر عضلانى موسوم به ناودان سیناپسى(synaptic gutter) و فضاى بین ترمینال عصبى و غشاء فیبر عضلانى موسوم به شکاف سیناپسى(synaptic cleft) است. در عمق ناودان سیناپسى، چین خوردگی هاى متعددى در فیبر عضلانى موسوم به شکاف هاى زیرعصبى(subneural clefts) وجود دارد. در ترمینال آکسون تعداد زیادى میتوکندرى وجود دارد که انرژى لازم براى سنتز میانجى تحریکى استیل کولین را تأمین مى کنند.استیل کولین در سیتوپلاسم ترمینال آکسونى ساخته مى شود اما به سرعت جذب تعداد زیادى وزیکول هاى سیناپسى کوچک مى گردد.تقریبا ۳۰۰۰۰۰ وزیکول سیناپسى به طور طبیعى در تمام ترمینال هاى یک صفحه انتهایى وجود دارند. بر روى سطح چین خوردگی هاى ناودان سیناپسى دستجاتى از آنزیم کولین استراز قرار دارد که قادر به تجزیه استیل کولین است.
در تصویر ذیل نماهای متفاوت صفحه حرکتی انتهایی را می بینید:
 هنگامی که یک ایمپالس عصبى به محل تماس عصبى- عضلانى مى رسد حدود ۳۰۰ وزیکول استیل کولین به وسیله ترمینال ها به داخل شکاف هاى سیناپسى بین ترمینال ها و غشاء فیبر عضلانى آزاد مى شوند. این امر ناشى از حرکت یون هاى کلسیم از مایع خارج سلولى به داخل غشاء ترمینال ها در هنگامى است که پتانسیل عمل غشاء ترمینال ها را دپولاریزه مى کند. یون هاى کلسیم موجب پاره شدن وزیکول هاى استیل کولین به داخل شکاف سیناپسى مى شوند(شکل زیر). در غیاب یون کلسیم یا در حضور مازاد منیزیوم، آزاد شدن استیل کولین شدیدا تضعیف مى شود.
هنگامی که یک ایمپالس عصبى به محل تماس عصبى- عضلانى مى رسد حدود ۳۰۰ وزیکول استیل کولین به وسیله ترمینال ها به داخل شکاف هاى سیناپسى بین ترمینال ها و غشاء فیبر عضلانى آزاد مى شوند. این امر ناشى از حرکت یون هاى کلسیم از مایع خارج سلولى به داخل غشاء ترمینال ها در هنگامى است که پتانسیل عمل غشاء ترمینال ها را دپولاریزه مى کند. یون هاى کلسیم موجب پاره شدن وزیکول هاى استیل کولین به داخل شکاف سیناپسى مى شوند(شکل زیر). در غیاب یون کلسیم یا در حضور مازاد منیزیوم، آزاد شدن استیل کولین شدیدا تضعیف مى شود.
 با وجود اینکه استیل کولین آزاد شده به داخل شکاف بین صفحه انتهایى و غشاء عضلانى فقط براى جزء بسیار کوچکى از یک ثانیه در آنجا باقى مى ماند، در همین زمان کوتاه نفوذپذیرى غشاء عضله به یون هاى مثبت چندین هزار برابر افزایش مى یابد که علت این امر باز شدن کانالهاى یونى دریچه دار وابسته به استیل کولین است چون استیل کولین به نوبه خود موجب یک تغییر شکل فضایى در مولکول کانال دار مى شود که دریچه آن را براى حدود یک میلى سکند باز مى کند.
با وجود اینکه استیل کولین آزاد شده به داخل شکاف بین صفحه انتهایى و غشاء عضلانى فقط براى جزء بسیار کوچکى از یک ثانیه در آنجا باقى مى ماند، در همین زمان کوتاه نفوذپذیرى غشاء عضله به یون هاى مثبت چندین هزار برابر افزایش مى یابد که علت این امر باز شدن کانالهاى یونى دریچه دار وابسته به استیل کولین است چون استیل کولین به نوبه خود موجب یک تغییر شکل فضایى در مولکول کانال دار مى شود که دریچه آن را براى حدود یک میلى سکند باز مى کند.
کانال استیل کولین قطرى حدود شصت و پنج صدم نانومتر دارد یعنى آنقدر بزرگ است که به تمام یون هاى مثبت مهم نظیر سدیم، پتاسیم و کلسیم اجازه مى دهد تا به آسانى از کانال عبور کنند. یون هاى منفى از قبیل یون هاى کلر از این کانال عبور نمى کنند و علت این امر وجود بارهاى منفى قوى در جدار کانال است.
ریختن ناگهانى یون هاى سدیم به داخل فیبر عضلانى در هنگام باز بودن کانال هاى استیل کولینى موجب مى شود که پتانسیل غشاء در ناحیه صفحه انتهایى در جهت مثبت به میزان ۵۰ تا ۷۰ میلى ولت افزایش یابد و یک پتانسیل موضعى موسوم به پتانسیل صفحه انتهایى تولید کند.
با توجه به این مسأله که یک افزایش ناگهانى در پتانسیل غشاء به میزانى بیش از ۱۵ تا ۳۰ میلى ولت براى شروع مکانیسم فیدبکى مثبت فعال شدن کانالهاى سدیمى کافى است مى توانیم درک کنیم که پتانسیل صفحه انتهایى ایجاد شده توسط تحریک استیل کولین بسیار بیشتر از حد کافى براى تولید پتانسیل عمل در فیبر عضلانى است.
در حالت استراحت، نیروهاى جاذبه اى بین فیلامان هاى اکتین و میوزین مهار مى شوند اما هنگامى که یک پتانسیل عمل در غشاء فیبر عضلانى سیر مى کند موجب آزاد شدن مقدار زیادى یون کلسیم توسط رتیکولوم سارکوپلاسمیک به داخل سارکوپلاسم (سیتوپلاسم فیبر عضلانى) اطراف میوفیبریل ها مى گردد. این یون هاى کلسیم، نیروهاى جاذبه اى بین فیلامان هاى اکتین و میوزین را فعال مى کنند و انقباض فیبر عضلانى شروع مى شود (تصویر زیر).
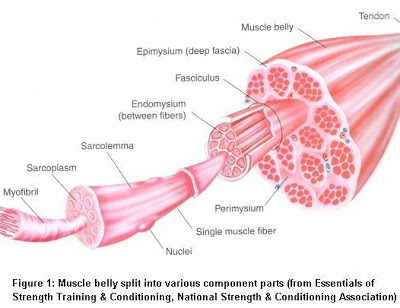
 همانطور که مى دانیم هر عضله اسکلتی از تعداد زیادی فیبر عضلانى که هریک از این فیبرها خود محتوى صدها تا هزارها میوفیبریل است تشکیل می گردد و هر میوفیبریل نیز به نوبه خود داراى حدود ۱۵۰۰ فیلامان میوزین و دو برابر آن فیلامان هاى اکتین است که در کنار یکدیگر قرار گرفته اند(تصاویر ذیل و فوق).
همانطور که مى دانیم هر عضله اسکلتی از تعداد زیادی فیبر عضلانى که هریک از این فیبرها خود محتوى صدها تا هزارها میوفیبریل است تشکیل می گردد و هر میوفیبریل نیز به نوبه خود داراى حدود ۱۵۰۰ فیلامان میوزین و دو برابر آن فیلامان هاى اکتین است که در کنار یکدیگر قرار گرفته اند(تصاویر ذیل و فوق).
 درحقیقت هر میوفیبریل از یک سری واحدهای تکراری بنام سارکومر تشکیل یافته است که واحد عملکردی انقباض عضله می باشد که خود هر سارکومر دارای فیلامان های میوزین و اکتین می باشد که کنار یکدیگر قرار گرفته اند به طوریکه اکتین در طرفین و میوزین در وسط واقع می شود.هرسارکومر بین دونوار Z واقع می شود که خط Z یک شبکه پروتئینی اتصالی است.
درحقیقت هر میوفیبریل از یک سری واحدهای تکراری بنام سارکومر تشکیل یافته است که واحد عملکردی انقباض عضله می باشد که خود هر سارکومر دارای فیلامان های میوزین و اکتین می باشد که کنار یکدیگر قرار گرفته اند به طوریکه اکتین در طرفین و میوزین در وسط واقع می شود.هرسارکومر بین دونوار Z واقع می شود که خط Z یک شبکه پروتئینی اتصالی است.
مولکول های میوزین از انتها به هم متصل بوده و باند تیره وسیع A را ایجاد می کنند.باند I و زون H قسمت هایی هستند که در آن نواحی بین میوفیلامان های اکتین و میوزین هم پوشانی وجود ندارد.باند I فقط دارای مولکول های ظریف اکتین و زون H فقط دارای میوفیلامان های میوزین می باشد.در وسط زون H هم خط M قرار گرفته است که این خط توسط پروتئین هایی به وجود می آید که تمام فیلامان های ضخیم میوزین را به یکدیگر متصل می سازد.
اینها مولکول هاى پروتئینى پلیمریزه بزرگى هستند که مسئول انقباض عضله مى باشند. فیلامانهاى ضخیم از جنس میوزین و فیلامانهاى نازک از جنس اکتین هستند. در ظرف تقریبا یک میلى سکند بعد از آنکه استیل کولین توسط ترمینال آکسونى آزاد شد قسمت زیادى از آن به خارج از شکاف سیناپسى انتشار یافته و دیگر بر روى غشاء فیبر عضلانى عمل نمى کند و قسمت باقیمانده توسط کولین استراز موجود در تیغه هاى پایه بین ترمینال عصبى و شکاف هاى زیرعصبى منهدم مى شود. پس از هیدرولیز، کولین با انتقال فعال به آکسوپلاسم باز مى گردد. دوره زمانى بسیار کوتاهى که استیل کولین در تماس با غشاء فیبر عضلانى باقى مى ماند ــ حدود یک میلى سکند ــ تقریبا همیشه براى تحریک فیبر عضلانى کافى است و به علاوه، حذف سریع استیل کولین، بعد از آنکه اثر پتانسیل عمل اول بر روى فیبر عضلانى از بین رفت، از تحریک مجدد فیبر عضلانى جلوگیرى مى کند.
به نقل از فیزیوتراپی (Physical Therapy )
